Ratusnik Test 1 Focus Review Neuro Basis of Speech and Communication
Introduction
Conversation involves an extremely complicated set of processes in which participants have to interweave their activities with precise timing, and notwithstanding information technology is a skill that all speakers seem very practiced at (Garrod and Pickering, 2004). Ane argument for why conversation is so easy is that interlocutors tend to become aligned at different levels of linguistic representation and therefore discover it easier to perform this joint activity than the individual activities of speaking or listening (Garrod and Pickering, 2009). Pickering and Garrod (2004) explain the process of alignment in terms of their interactive-alignment business relationship. According to this account, conversation is successful to the extent that participants come to understand the relevant aspects of what they are talking about in the aforementioned mode. More than specifically, they construct mental models of the situation under give-and-take (i.e., situation models; Zwaan and Radvansky, 1998), and successful chat occurs when these models get aligned. Interlocutors usually practice not align deliberately. Rather, alignment is largely the outcome of the tendency for interlocutors to repeat each other's linguistic choices at many different levels, such as words and grammar (Garrod and Anderson, 1987; Brennan and Clark, 1996; Branigan et al., 2000). Such alignment is, therefore, a form of fake. Essentially, interlocutors prime each other to speak most things in the aforementioned way, and people who speak about things in the same way tend to think about them in the aforementioned way likewise.
At the level of state of affairs models, interlocutors align on spatial reference frames: if one speaker refers to objects egocentrically (e.g., "on the left" to mean on the speaker'south left), then the other speaker tends to use an egoistic perspective besides (Watson et al., 2004). More by and large, they align on a characterization of the representational domain, for case using coordinate systems (due east.g., A4, D3) or figural descriptions (e.g., T-shape, right indicator) to refer to positions in a maze (Garrod and Anderson, 1987; Garrod and Doherty, 1994). They also echo each other'southward referring expressions, even when they are unnecessarily specific (Brennan and Clark, 1996). Faux also occurs for grammer, with speakers repeating the syntactic structure used past their interlocutors for cards describing events (Branigan et al., 2000; east.yard., "the diver giving the block to the cricketer") or objects (Cleland and Pickering, 2003; e.thousand., "the sheep that is red"), and repeating syntax or closed-form lexical items in question-answering (Levelt and Kelter, 1982). Bilinguals even repeat syntax betwixt languages, for example when ane interlocutor speaks English language and the other speaks Spanish (Hartsuiker et al., 2004). Finally, in that location is prove for alignment of phonetics (Pardo, 2006), and of accent and speech charge per unit (Giles et al., 1991).
An of import property of interactive alignment is that it is automatic in the sense that speakers are non aware of the process and that it does not appear effortful. Such automatic imitation or mimicry occurs in social situations more by and large. Thus, Dijksterhuis and Bargh (2001) argued that many social behaviors are automatically triggered by perception of the deportment of other people, in a way that frequently leads to simulated (eastward.g., Chartrand and Bargh, 1999). We propose that the automatic alignment channels linking unlike levels of linguistic representation operate in essentially the same mode (encounter Figure 1). In other words, conversationalists do not demand to decide to interpret each different level of linguistic representation for alignment to occur at all these channels (Pickering and Garrod, 2006). This is considering the alignment channels reflect priming rather than deliberative processing. In add-on there are aspects of automatic not-linguistic imitation that can facilitate alignment at linguistic levels (Garrod and Pickering, 2009). For example, when speakers and listeners align their gaze to look at the same affair this can facilitate alignment of interpretation (Richardson and Dale, 2005; Richardson et al., 2007). The opposite as well appears to hold, with linguistic alignment enhancing romantic allure, which presumably involves not-linguistic alignment (Ireland et al., 2011).
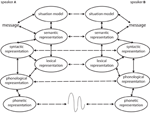
Figure 1. The interactive-alignment model (based on Pickering and Garrod, 2004). Speakers A and B represent two interlocutors in a dialogue in this schematic representation of the stages of comprehension and product co-ordinate to the model. The dashed lines correspond alignment channels.
The interactive alignment account makes two basic assumptions nearly language processing in dialogue. First, there is parity of representations used in speaking and listening. The same representations are used during production (when speaking) and comprehension (when listening to some other person). This explains why linguistic repetition occurred in experiments such equally Branigan et al. (2000), who had participants have turns to describe and match picture cards, and found that they tended to use the form of utterance just used by their partner. For case, they tended to use a "prepositional object" class such as the pirate giving the book to the swimmer following another prepositional object sentence but a "double object" form such as the pirate giving the swimmer the volume following some other double object sentence (though both sentences have essentially the same meaning). In such cases, the aforementioned grammatical representation is activated during speaking and listening. For a different class of testify for syntactic parity, see Kempen et al. (2011).
Second, the processes of alignment at dissimilar levels (e.yard., words, construction, meaning) interact in such a manner that increased alignment at one level leads to increased alignment at other levels (i.east., alignment percolates betwixt levels). In this review, we examine the neural evidence for these ii assumptions. For example, alignment of syntactic structure is enhanced by repetition of words, with participants existence even more likely to say The cowboy handing the assistant to the infiltrator afterwards hearing The chef handing the jug to the swimmer than afterward The chef giving the jug to the swimmer (Branigan et al., 2000). Thus, alignment at one level (in this example, lexical alignment) enhances alignment at another level (in this example, grammatical alignment). Similarly, people are more likely to apply an unusual form such as the sheep that's scarlet (rather than the red sheep) after they take simply heard the goat that'southward ruddy than after they heard the door that's red (Cleland and Pickering, 2003). This is considering alignment at the semantic level (in this instance, with respect to animals) increases syntactic alignment. Furthermore, alignment of words leads to alignment of situation models—people who describe things the same style tend to think about them in the same fashion too (Markman and Makin, 1998). This means that alignment of low-level structure can eventually affect alignment at the crucial level of speakers' state of affairs models, the hallmark of successful communication.
In this review, nosotros appraise the neural evidence for the interactive alignment model. Nosotros focus on 3 central points. The first is that parity of representations exists between speaking and listening. This is a necessary, though not sufficient, condition for interactive alignment between interlocutors to be possible. The second is that alignment at one level of representation affects alignment at another. We review what evidence is available, and suggest concrete avenues for farther research. The tertiary is that alignment of representations should be related to mutual understanding. Further, we briefly explore how alignment between interlocutors may also play a office in controlling non-linguistic aspects of a conversation.
Neural Evidence
Testify for Parity
If interactive alignment of dissimilar linguistic representations betwixt speakers and listeners is to be possible, and so these representations demand to exist coded in the same form irrespective of whether a person is speaking or listening. There needs to be parity of representations between language comprehension and production. If this parity exists, then presumably, the neuronal infrastructure underlying the processing of language at different levels of representation should be the aforementioned during speaking and listening. This is a prerequisite for neural alignment during conversation, in which both interlocutors speak and heed. Neural parity underlies Hasson et al.'due south (2012) brain-to-brain coupling principle, in which parity emerges from the process by which the perceiver's perceptual system codes for an actor'south beliefs.
Below, we review the bear witness for parity of neural representations in speaking and listening across dissimilar linguistic levels. We focus mainly on studies that take either directly compared the two modalities, or manipulated 1 while observing the other. The number of relevant studies is limited because neuroimaging evidence on linguistic communication production is much scarcer than on language comprehension. Many of the studies, in item those concerned with higher-level processes, investigate whether different modalities engage the same brain regions. This comparison yields less-than-perfect evidence for parity, because information technology is possible that the aforementioned encephalon region might code unlike representations, just it does provide suggestive evidence.
Perception and production of speech sounds
Much of the debate on the neuronal overlap between activeness and perception in language has focused on the role of the motor system. In their motor theory of speech perception, Liberman and Mattingly (1985) proposed that perceiving spoken language is to perceive the articulatory gestures one would make to produce the same speech. Thank you largely to the discovery of mirror neurons (Rizzolatti and Arbib, 1998), this theory has received renewed interest (Galantucci et al., 2006). It receives back up from TMS studies that have shown that listening to speech affects the excitability of brain regions controlling articulatory muscles (Watkins et al., 2003; Pulvermüller et al., 2006; Watkins and Paus, 2006). fMRI has provided converging show for enhanced motor cortex activity when listening to spoken language compared to balance (Wilson et al., 2004).
These studies show motor cortex involvement in perceiving speech, just they do not make articulate the verbal role of the motor cortex. According to motor theory, the main motor cortex activity should be specific to the sounds perceived. According to a proposal past Scott et al. (2009), motor cortex involvement in speech perception could instead reverberate a process general to the act of perceiving speech. In this proposal, the motor involvement reflects a readiness on the part of the listener to take part in the conversation and hence embark speaking. However, TMS studies suggest that the motor cortex activeness during speech perception is in fact specific to the sounds being articulated, as motor theory would predict: the excitability of articulators through TMS to the primary motor cortex is stronger when perceiving sounds that crave those articulators than when perceiving sounds that require different articulators (Fadiga et al., 2002; D'Ausilio et al., 2009). These claims are further supported by recent behavioral bear witness that the interference from distractor words on articulation is greater when the distractors contain sounds incompatible with the joint target (Yuen et al., 2010).
In improver, primary motor cortex response to videos of words being uttered depends on the articulatory complication of these words (Tremblay and Small, 2011). This suggests that the motor interest when listening to speech is related to the effort required to produce the same speech, once again suggesting that motor cortex involvement in voice communication perception is specific to the content of the perceived spoken language. A different measure of articulatory attempt, sentence length, fails to support this observation: when listening to sentences, principal motor cortex response does not appear to depend on the length of sentences being heard (Menenti et al., 2011; also see beneath). Hence, it is possible that the effect of articulatory effort on motor involvement in speech perception is specific to observing videos, or that it is somehow observable when listening to single words simply not when listening to sentences. In this context, it is worth noting that many of the studies showing motor involvement in perception use highly artificial paradigms (e.g., presenting phonemes in isolation or degrading the stimulus), and often compare speech to radically different, often less complex, acoustic stimuli, so it is possible that motor effects in natural spoken language perception could exist less pronounced (McGettigan et al., 2010). The aught finding in a study studying motor interest in more than naturalistic spoken communication perception (Menenti et al., 2011) could exist an indication in this direction.
Now that there is clear evidence for some motor involvement in speech perception, the debate has shifted to whether this interest is a necessary component of perceiving oral communication. Researchers from the mirror neuron tradition argue for a causal role of the motor cortex involvement in spoken communication perception described above, much along the lines of motor theory of speech perception (Pulvermüller and Fadiga, 2010). Merely an alternative view proposes that motor activation can occur, but that it is non necessary. The involvement has been characterized as modulatory (Hickok, 2009; Lotto et al., 2009; simply see Wilson, 2009) or equally beingness specific to certain situations and materials (Toni et al., 2008). In any case, show showing a link between specific backdrop of speech sounds being perceived and the articulators needed to produce them suggest that there is a link between representations.
In summary, there is considerable show for neural parity at the level of oral communication sounds. In contrast, the evidence for neural parity at higher linguistic levels is much scarcer. In detail, the technical difficulties associated with investigating speaking in fMRI increment as the stimuli go longer (eastward.grand., words, sentences, narratives). Furthermore, psycholinguistics (dissimilar work on the articulation and perception of oral communication) has generally assumed that comprehension and production of language have little to practise with each other. We now review what is available on lexical and syntactic processing in plough.
Parity of lexical processing
For processing of words, two similar studies contrasted processing of intransitive (e.g., jump) and transitive (e.1000., hitting) verbs in either voice communication product (Den Ouden et al., 2009) or comprehension (Thompson et al., 2007). In the production written report, the verbs were elicited using pictures or videos, and in comprehension the subjects read the verbs. The two studies produced very different results for the 2 modalities: in the comprehension study (Thompson et al., 2007), only one cluster in the left inferior parietal lobe showed a significant difference between the two kinds of verb. Despite the fact that a much larger distributed network of areas showed the consequence in production (Den Ouden et al., 2009), that one cluster was not part of the product network.
However, studies that straight compare production with comprehension or dispense the ane while investigating the other do notice that words share neural processes between the two modalities. In an intra-operative mapping report, Ilmberger et al. (2001) directly stimulated the cortex during comprehension and production of words. The ii tasks used had previously been shown to share a lot of variance, which was taken to indicate that they tapped into similar processes. Twelve out of xiv patients had sites where stimulation afflicted both naming and comprehension performance. Many of these sites were in left inferior frontal gyrus. This region contains Brodmann area (BA) 44, which has been shown to be involved both in lexical selection in speaking and in lexical decision in listening (Heim et al., 2007, 2009). Menenti et al. (2011) reported an fMRI adaptation study that compared semantic, lexical, and syntactic processes in speaking and listening. They institute that repetition of lexical content across heard or spoken sentences induced suppression effects in the same fix of areas (left anterior and posterior middle temporal gyrus and left inferior frontal gyrus) in both speaking and listening, although the precuneus additionally showed an adaptation effect in speaking but non listening. On the whole, then, there seems to be some evidence that the linguistic processing of words is accomplished by similar brain regions in speaking and listening.
Parity of syntax
In that location is somewhat clearer evidence for neural parity of syntax. Such work builds on theoretical and behavioral studies that support parity of syntactic representations (Branigan et al., 2000). Heim reviewed fMRI data on processing syntactic gender and concluded that speaking and listening rely on the aforementioned network of encephalon areas, in particular BA 44 (Heim, 2008). In addition, Menenti et al.'s (2011) fMRI accommodation study showed that repetition of syntactic structure (as found in agile and passive transitive sentences) induced suppression effects in the same brain regions (BA 44 and BA 21) for speaking and listening. However, in a PET study on comprehension and production of syntactic structure, Indefrey and colleagues found effects of syntactic complexity in speech production (in BA 44), only non comprehension (Indefrey et al., 2004). They interpreted their data in terms of theoretical accounts in which listeners need non always fully encode syntactic structure merely can instead rely on other cues to empathise what is existence said (Ferreira et al., 2002), but where speakers e'er construct complete syntactic representations. However, it is also possible that this lack of parity is due to chore requirements rather than indicating general differences between product and comprehension.
Importantly, as mentioned in a higher place, studies showing that the same encephalon regions are involved in two modalities do not prove that the same representations or even the aforementioned processes are being recruited. Feasibly, different neuronal populations in the aforementioned full general encephalon regions could process syntax in speaking and listening respectively. To address this issue, Segaert et al. (2011) used the same paradigm as Menenti et al. (2011), but this fourth dimension intermixing comprehension and product trials within the same experiment. Participants therefore produced or heard transitive sentences in interspersed order and the syntactic construction of these sentences could exist either novel or repeated across sentences. This produced cantankerous-modal adaptation furnishings and no interaction between the size of the issue and whether priming was intra- or inter-modal. This strongly supports the idea that the same neuronal populations are being recruited for the production and comprehension of syntax in speaking and listening, and hence that the neural representations involved in the two modalities are alike.
So far nosotros accept reviewed evidence for parity of different types of linguistic representations in speaking and listening, only in intra-private settings. While such parity is a necessary condition for alignment, it is not a sufficient one: a central tenet of interactive alignment is that representations get more aligned over the course of dialogue. Testing this tenet requires studies in which bodily between-participant communication takes place, and in which different levels of representation can be segregated in terms of their neural signature. These studies, unfortunately, yet need to exist done.
Percolation
The interactive alignment business relationship farther predicts that alignment at i level of representation leads to alignment at other levels of representation too. To exam this prediction, it is necessary to conduct studies in chatty settings that somehow target at least ii levels of representation. In the introduction, we have noted behavioral evidence from structural priming (Cleland and Pickering, 2003). In another behavioral study, Adank et al. (2010) showed that alignment of spoken communication sounds can improve comprehension. Participants were tested on their comprehension of sentences in an unfamiliar emphasis presented in noise. They then underwent ane of several types of training: no training; just listening to the sentences; transcribing the sentences; repeating the sentences in their ain accent; repeating the sentences while imitating the accent; and doing so in noise so that they could not hear themselves speak. They were then tested on comprehension for a dissimilar set of similar sentences. But the 2 imitation conditions improved comprehension performance in the post-exam. This suggests that assuasive a listener to align with the speaker at the audio-based level that is required to produce the output improves comprehension.
In a study investigating gestural communication, Schippers and colleagues scanned pairs of players in a game of charades (Schippers et al., 2010). They beginning scanned the gesturer and videotaped his or her gestures, and and then they scanned the guesser while he or she was watching the videotape. Using Granger Causality Mapping, they looked for encephalon regions whose action in the gesturer predicted that in the guesser. Starting time, they establish Granger-causation betwixt the "putative mirror neuron system (pMNS)"—defined equally dorsal and ventral premotor, somatosensory cortex, anterior inferior parietal lobule, and midtemporal gyrus—from the gesturer to the guesser. This provides further support for the extensive literature arguing for overlap in neural processes between action and perception (Hasson et al., 2012). In addition, they establish Granger-causation between the gesturer'due south pMNS and the guesser's ventromedial prefrontal cortex, an area that is involved in inferring someone'due south intention (i.e., mentalizing; Amodio and Frith, 2006). Dale and colleagues used the tangram task, a dialogue task known to elicit progressively more similar lexical representations from interlocutors, to bear witness that over time interlocutors' heart movements also go highly synchronous (Dale et al., 2011). Alignment in lexical representation here, therefore, co-occurs with alignment in behavior. Farther, Broca'southward area has frequently been found involved in both producing and comprehending language at diverse levels (Bookheimer, 2002; Hagoort, 2005), and in producing and comprehending actions (Rossi et al., 2011), suggesting a potential neural substrate for percolation between these ii levels of representation. Together, these data propose that alignment between chat partners occurs from lower to higher levels of representation, and also between non-linguistic and linguistic processes.
Absolutely, neural evidence for (or against) percolation is deficient. Equally mentioned in the introduction, the lexical boost in syntactic priming is ane example of percolation. This lexical boost could exist used in an fMRI report past comparing syntactic priming between interlocutors in conditions with and without lexical repetition. For example, if the study past Menenti et al. (2011) was repeated in an interactive setting, then the extent of lexical repetition suppression across participants should correlate with syntactic priming. If inter-bailiwick correlations in encephalon activeness reflect alignment (Stephens et al., 2010; see beneath), alignment at i level (e.k., sound) could be manipulated, and the extent of correlation betwixt subjects also equally comprehension could be assessed. Phonological alignment should impact the inter-subject correlations, and in item, it should affect those inter-subject area correlations that also correlate with the comprehension score of the subject.
Ultimate Goal of Communication: Alignment of Situation Models
According to the interactive alignment business relationship, conversation is successful to the extent that participants come to empathise the relevant aspects of what they are talking about in the aforementioned way. Ultimately, therefore, alignment of situation models is crucial—both to advice and to the interactive alignment account.
In an fMRI report, Awad et al. (2007) showed that a similar network of areas is involved in comprehending and producing narrative speech. Nevertheless, production and comprehension were each contrasted with radically different baseline atmospheric condition before being compared to one another, making the results hard to interpret. In their fMRI adaptation study on overlap between speaking and listening, Menenti et al. (2011) likewise looked at repetition of sentence-level significant. As for lexical repetition and syntactic repetition, they institute that the aforementioned brain regions (in this case, the bilateral temporoparietal junction) show adaptation effects irrespective of whether people are speaking or listening, suggesting a neuronal correlate for parity of meaning. This study, nevertheless, left unanswered the question at which level of pregnant parity of representations held: was it the not-exact state of affairs model underlying the sentences, or the linguistic meaning of the sentences itself?
In a follow-upward written report on sentence product, Menenti et al. (2012) thus further distinguished between repetition of the linguistic meaning of sentences (the sense) or the underlying mental representation (the reference). For example, if the sentence The man kisses the adult female was used twice to refer to dissimilar subsequent pictures, this constituted a repetition of sense. Conversely, the same motion-picture show of a homo kissing a adult female could be shown outset with the sentence The red human kisses the green woman and then with the judgement The yellowish man kisses the blue adult female, leading to a repetition of reference. The brain regions previously shown to have similar semantic repetition effects in speaking and listening (Menenti et al., 2011) turned out to be mainly sensitive to repetition of referential meaning: they showed suppression effects when the same picture was repeated even if with a dissimilar sentence, but did not showroom any such sensitivity to the repetition of sentences themselves if accompanied by dissimilar pictures. This suggests alignment of underlying non-linguistic representations in speaking and listening, rather than purely alignment of linguistic semantic structure.
It is also possible to investigate alignment of significant in a more naturalistic manner, while still allowing for a detailed analysis. The drawback of naturalistic experiments is oftentimes that interpretations are hard to depict because the relevant details of the stimulus are non clear. This problem can be circumvented past using subjects equally models for each other, an inter-subject correlation approach (Hasson et al., 2004). The idea is that if there are areas where subjects' brain activity is the same over the whole time-class of a stimulus, these correlations in encephalon activity are likely to be driven by that stimulus, whatsoever the stimulus may be. Stephens et al. (2010) used this approach to investigate inter-subject correlations in fMRI between a speaker and a group of listeners. They first recorded a speaker in the scanner while she was telling an unrehearsed story, then recorded listeners who heard that story. Correlations between speakers and listeners occurred in many different brain regions. These correlations were positively related to listeners' comprehension (as measured by a subsequent examination). When a grouping of listeners were presented a story in an unfamiliar linguistic communication (Russian), these correlations disappeared. This suggests that alignment in brain action between a speaker communicating information and a listener hearing it is tied to the agreement of that information.
In a study on listeners only, Lerner et al. (2011) studied inter-subject correlations for four levels of temporal structure: reversed spoken communication, a word list, a listing of paragraphs, and a story. They found that equally the temporal construction of the materials increased (i.due east., they were closer to complete stories), the correlations between participants extended from auditory cortex further posterior and into the parietal lobes. This report was conducted with listeners just, and thus did not properly target alignment betwixt interlocutors in communication. Nonetheless it provides indirect evidence: the interactive alignment business relationship assumes that listeners align with speakers. Different listeners of the same speaker should, therefore, likewise align. Building on the speaker-listener correlations shown by Stephens et al. (2010) listener-listener correlations tin, so, tell u.s.a. something near neural alignment. These findings provide some evidence that alignment at several levels of representation leads to more than all-encompassing correlations in brain activity. Even so, for both Stephens et al. (2010) and Lerner et al. (2011) a discussion of circumspection is in order: both studies showed an effect (in this case, a correlation) in one condition but not the other (in Stephens et al., dissimilar languages; in Lerner et al. different temporal structures); they did not show that the conditions were significantly different.
These studies provide evidence that situation models for fifty-fifty very complex stimuli tin be usefully investigated past using novel analysis techniques. They propose that alignment can be tracked in the brain, and can be measured in time too (Hasson et al., 2012). More than piece of work is needed though: while these studies suggest that alignment can be operationalized as inter-subject area correlations, and that these are related to understanding, dissimilar levels of representations can only be distinguished indirectly, by mapping the findings onto other studies that haven't necessarily targeted communication. An important avenue for further enquiry, therefore, is to investigate in more detail to what aspects of communication correlations in different brain regions are due. Furthermore, the interactive alignment account assumes that dialogue is not just an expanded monologue. Therefore, if we want to notice out how dialogue works, we will demand to get and study dialogue.
Not-Linguistic Aspects of Dialogue
The interactive-alignment model assumes that successful communication is supported by interlocutors aligning at many different levels of representation. Above, we have reviewed studies concerned with linguistic representations. Just language alone is non sufficient to have a proper conversation (Enrici et al., 2010; Willems and Varley, 2010). Alignment between interlocutors may likewise exist occurring for boosted non-linguistic processes that are necessary to keep a conversation flowing. In Section "Ultimate Goal of Communication: Alignment of Situation Models" we discussed a few examples of where alignment of not-linguistic processes may percolate into alignment of linguistic representations. Beneath, we bear upon upon proposals of how alignment of not-linguistic processes may help govern the act of holding a chat.
During conversation, we do non only use language to convey our intentions. Torso posture, prosody, and gesture are vital aspects of conversation and are taken into account effortlessly when trying to infer what a speaker intends. Abundant evidence suggests that gesture and speech comprehension and production are closely related (Willems and Hagoort, 2007; Enrici et al., 2012). Percolation betwixt gesture and spoken communication could, therefore, occur just like percolation within levels of representation in speech. The extensive literature on the mirror neuron system shows that activity ascertainment and activity execution are intimately intertwined (Fabbri-Destro and Rizzolatti, 2008), suggesting a plausible neural correlate for alignment at the gestural level between interlocutors. Communicative gestures take indeed been shown to produce related encephalon activity in observers' and gesturers' pMNSs (Schippers et al., 2010).
One time a person has settled on a message, they may demand to decide how all-time to convey it in a particular setting to a particular partner. A prepare of studies targeted the generation or recognition of such chatty intentions in verbal and non-verbal advice. Both tasks were designed to make advice difficult and hence heighten the demand for such processes: in the not-verbal experiment, participants devised a novel form of communication using only the movement of shapes in a grid (Noordzij et al., 2009). In the exact experiment, participants described words to each other, but were not allowed to use words highly associated with the target (Willems et al., 2010). Both studies showed that sending and receiving these letters involved the same brain region: the correct temporo-parietal junction in non-verbal advice, and the left temporo-parietal junction in verbal communication. These studies support parity for chatty processes in verbal and not-verbal communication, respectively. However in neither study was feedback allowed, so they would need to be generalized to interactive dialogue.
Another important aspect of holding a smooth conversation is turn taking. While we may well be attuned to what our partner intends to say, if we fail to rails when it is our turn to speak and when information technology is not, then we are likely to suspension excessively between contributions, speak at the aforementioned time, or interrupt each other, leaving the conversation with niggling chance of success. 1 account has alignment of neural oscillations playing a major role in conversation (Wilson and Wilson, 2005). In this account, the production organisation of a speaker oscillates with a syllabic stage: the readiness to initiate a new syllable is at a minimum in the middle of a syllable and peaks half a cycle after syllable outset. Conversation partners' oscillation rates get entrained over the class of a chat, but they are in anti-phase, so that the speaker'south readiness to speak is at minimum when the listener'southward is at a maximum, and vice versa (Gambi and Pickering, 2011; Hasson et al., 2012). A farther hypothesis is that the theta frequency range is primal to this machinery: across languages, typical spoken language product is three–8 syllables per 2d (Drullman, 1995; Greenberg et al., 2003; Chandrasekaran et al., 2009). Auditory cortex has been shown to produce ongoing oscillations at this frequency (Buzsaki and Draguhn, 2004). A possibility is that the ongoing oscillations resonate with the incoming voice communication at the same frequency, thereby amplifying the betoken. This means that the neural oscillations in the theta frequency band become entrained between listeners and speakers, and that this aids advice (Hasson et al., 2012).
Entrainment at the syllable frequency, however, cannot be enough to explain turn-taking as we don't normally want to interrupt our interlocutors at every syllable (Gambi and Pickering, 2011). Recently, Bourguignon et al. (2012) demonstrated coherence between a speakers' speech product (f0 formant) and listener'southward brain oscillations around 0.5 Hz. This frequency is related to the prosodic envelope of voice communication. Indeed, the coherence was also present for unintelligible spoken language stimuli (a foreign language or a hummed text), but in different brain regions. Possibly, and so, resonating with our interlocutor's speech patterns at unlike frequencies enables us to meliorate predict when his turn will end (Giraud and Poeppel, 2012).
Future Directions
In the in a higher place, we have reviewed neural prove relevant to the interactive alignment model of chat. While neuroimaging studies on speech communication production of annihilation more circuitous than a single phoneme are still too scarce to provide a definite answer, the testify is mounting that speakers and listeners generally apply the same brain regions for the same types of stimuli. Indeed, when communicating, speakers' and listeners likewise show correlated brain activity. Alignment is, therefore, both possible and real.
But does neural alignment occur during interactive language? It would surely exist surprising if neural alignment occurred when speakers and listeners were separated, merely did not occur when they interacted (in function considering psycholinguistic evidence for alignment is based on dialogue; Pickering and Garrod, 2004). Yet, the current literature does not all the same directly answer this question. The field needs strategies to meaningfully report interacting participants. Promising approaches take been devised for non-linguistic live interaction (Newman-Norlund et al., 2007, 2008; Dumas et al., 2010, 2011; Redcay et al., 2010; Baess et al., 2012; Guionnet et al., 2012). It is fourth dimension that neuroimaging inquiry on language follows suit—non an easy challenge, as the dearth of studies attempting this shows. Technical challenges aren't the only issue when wanting to study conversation: with so little control over a stimulus; it is hard to devise experiments that provide precise and meaningful data. Gambi and Pickering (2011) provide suggestions for possible paradigms to written report interactive language use; these may as well be benign to neuroimaging inquiry on the topic.
Equally the attention of neuroscience turns toward the role of prediction (Friston and Kiebel, 2009; Friston, 2010; Clark, in press), interactive alignment provides a natural mechanistic basis on which predictions can be built. Pickering and Garrod (in press) propose a "simulation" business relationship in which comprehenders covertly imitate speakers and use those representations to predict upcoming utterances (and therefore gear up their own contributions accordingly). Comprehenders are more likely to predict appropriately when they are well-aligned with speakers; just in improver, the process of covert imitation provides a mechanism for alignment. This account assumes that production processes are used during comprehension (and in fact that comprehension processes are used during production).
Based on the interactive alignment model, we make the following predictions for further research on dialogue: (one) Alignment: speaking and listening make employ of similar representations, and hence have largely overlapping neural correlates. We accept reviewed the bachelor show for this prediction to a higher place, but more work is needed, particularly studies targeting both speaking and listening simultaneously: the overlap in neuronal correlates for each level of representation should further be increased in an interactive, communicative setting compared to not-communicative settings. (two) Percolation: alignment at one level of representation leads to alignment at other levels. In detail, alignment at lower levels of representation leads to better alignment of situation models, and thus, better communication. We have reviewed the (scarce) evidence available above, only truly putting this prediction to the test requires that studies of interacting interlocutors manipulate unlike levels of representation simultaneously, and furthermore have an outcome mensurate of communicative success. (3) Language processes are complemented by processes specific to a communicative setting. By carefully targeting both linguistic and non-linguistic aspects of conversation, future research will hopefully be able to demonstrate how these processes interact.
Conclusion
Nosotros have reviewed neural bear witness for the interactive alignment model of conversation. For linguistic processes, we have shown that representations in speaking and listening are similar, and that, hence, alignment between participants in a chat is at to the lowest degree possible. We have further reviewed evidence pertaining to the goal of a conversation, which is to communicate. As the interactive alignment model predicts, the ease of constructing a situation model is associated with increased correlation in brain activeness betwixt participants. Finally, we have touched upon literature dealing with alignment of processes more than specific to the act of communicating, and suggested how these might chronicle to the interactive-alignment model.
Conflict of Interest Statement
The authors declare that the research was conducted in the absenteeism of whatsoever commercial or financial relationships that could be construed equally a potential conflict of involvement.
References
Awad, M., Warren, J. E., Scott, S. Grand., Turkheimer, F. E., and Wise, R. J. S. (2007). A common system for the comprehension and product of narrative oral communication. J. Neurosci. 27, 11455–11464.
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Baess, P., Zhdanov, A., Mandel, A., Parkkonen, 50., Hirvenkari, L., Mäkelä, J. P., Jousmäki, V., and Hari, R. (2012). MEG dual scanning: a procedure to study existent-time auditory interaction betwixt two persons. Front. Hum. Neurosci. 6:83. doi: 10.3389/fnhum.2012.00083
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Bourguignon, One thousand., de Tiège, X., de Beeck, M. O., Ligot, North., Paquier, P., van Bogaert, P., Goldman, Southward., Hari, R., and Jousmäki, V. (2012). The stride of prosodic phrasing couples the listener'southward cortex to the reader's vocalism. Hum. Brain Mapp. doi: 10.1002/hbm.21442. [Epub ahead of print].
Pubmed Abstruse | Pubmed Full Text | CrossRef Total Text
Brennan, S. East., and Clark, H. H. (1996). Conceptual pacts and lexical choice in conversation. J. Exp. Psychol. Learn. Mem. Cogn. 22, 1482–1493.
Pubmed Abstract | Pubmed Full Text
Chandrasekaran, C., Trubanova, A., Stillittano, S., Caplier, A., and Ghazanfar, A. A. (2009). The natural statistics of audiovisual spoken language. PLoS Comput. Biol. 5:e1000436. doi: 10.1371/journal.pcbi.1000436
Pubmed Abstract | Pubmed Total Text | CrossRef Total Text
Chartrand, T. L., and Bargh, J. A. (1999). The chameleon effect: the perception – Äìbehavior link and social interaction. J. Pers. Soc. Psychol. 76, 893–910.
Pubmed Abstract | Pubmed Full Text
Clark, A. (in press). Whatsoever adjacent? Predictive brains, situated agents, and the future of cerebral science. Behav. Brain Sci.
Cleland, A. A., and Pickering, Thousand. J. (2003). The use of lexical and syntactic information in linguistic communication production: show from the priming of noun-phrase structure. J. Mem. Lang. 49, 214–230.
D'Ausilio, A., Pulvermüller, F., Salmas, P., Bufalari, I., Begliomini, C., and Fadiga, L. (2009). The motor somatotopy of speech perception. Curr. Biol. 19, 381–385.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Den Ouden, D.-B., Fix, Southward., Parrish, T. B., and Thompson, C. Thou. (2009). Argument construction effects in activity verb naming in static and dynamic weather. J. Neurolinguist. 22, 196–215.
Dijksterhuis, A., and Bargh, J. A. (2001). "The perception-behavior superhighway: automated effects of social perception on social behavior," in Advances in Experimental Social Psychology, ed P. Z. Mark (San Diego, CA: Bookish Press), ane–forty.
Dumas, Grand., Lachat, F., Martinerie, J., Nadel, J., and George, Northward. (2011). From social behaviour to brain synchronization: review and perspectives in hyperscanning. IRBM 32, 48–53.
Enrici, I., Adenzato, 1000., Cappa, S., Bara, B. Chiliad., and Tettamanti, G. (2010). Intention processing in communication: a common brain network for language and gestures. J. Cogn. Neurosci. 23, 2415–2431.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Enrici, I., Adenzato, K., Cappa, S., Bara, B. G., and Tettamanti, 1000. (2012). Intention processing in communication: a common encephalon network for language and gestures. J. Cogn. Neurosci. 23, 2415–2431.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Ferreira, F., Bailey, Thou. G. D., and Ferraro, 5. (2002). Good-enough representations in language comprehension. Curr. Dir. Psychol. Sci. xi, eleven–fifteen.
Galantucci, B., Fowler, C. A., and Turvey, K. T. (2006). The motor theory of speech perception reviewed. Psychon. Balderdash. Rev. 13, 361–377.
Pubmed Abstract | Pubmed Full Text
Garrod, Southward., and Doherty, Thou. (1994). Chat, co-ordination and convention: an empirical investigation of how groups institute linguistic conventions. Knowledge 53, 181–215.
Pubmed Abstract | Pubmed Full Text
Garrod, S., and Pickering, M. J. (2009). Joint action, interactive alignment, and dialog. Height. Cogn. Sci. 1, 292–304.
Giles, H., Coupland, North., and Coupland, J. (eds.). (1991). Contexts of Adaptation: Developments in Applied Sociolinguistics. Cambridge, MA: Cambridge University Press.
Greenberg, Due south., Carvey, H., Hitchcock, L., and Chang, S. (2003). Temporal properties of spontaneous speech – a syllable-centric perspective. J. Phon. 31, 465–485.
Guionnet, S., Nadel, J., Bertasi, Due east., Sperduti, 1000., Delaveau, P., and Fossati, P. (2012). Reciprocal imitation: toward a neural basis of social interaction. Cereb. Cortex 22, 971–978.
Pubmed Abstract | Pubmed Total Text | CrossRef Total Text
Hartsuiker, R. J., Pickering, M. J., and Veltkamp, E. (2004). Is syntax carve up or shared between languages? Cantankerous-linguistic syntactic priming in Spanish/English bilinguals. Psychol. Sci. xv, 409–414.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasson, U., Ghazanfar, A. A., Galantucci, B., Garrod, Southward., and Keysers, C. (2012). Brain-to-encephalon coupling: a mechanism for creating and sharing a social world. Trends Cogn. Sci. sixteen, 114–121.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasson, U., Nir, Y., Levy, I., Fuhrmann, G., and Malach, R. (2004). Intersubject synchronization of cortical activity during natural vision. Science 303, 1634–1640.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heim, S., Eickhoff, S., Friederici, A., and Amunts, Thou. (2009). Left cytoarchitectonic expanse 44 supports option in the mental lexicon during linguistic communication product. Brain Struct. Funct. 213, 441–456.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Heim, S., Eickhoff, S., Ischebeck, A., Supp, G., and Amunts, K. (2007). Modality-independent involvement of the left BA 44 during lexical conclusion making. Brain Struct. Funct. 212, 95–106.
Pubmed Abstruse | Pubmed Full Text | CrossRef Total Text
Ilmberger, J., Eisner, Due west., Schmid, U., and Reulen, H.-J. (2001). Operation in picture naming and word comprehension: evidence for common neuronal substrates from intraoperative linguistic communication mapping. Brain Lang. 76, 111–118.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Indefrey, P., Hellwig, F., Herzog, H., Seitz, R. J., and Hagoort, P. (2004). Neural responses to the product and comprehension of syntax in identical utterances. Brain Lang. 89, 312–319.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Ireland, Yard. E., Slatcher, R. B., Eastwick, P. West., Pair of scissors, L. East., Finkel, E. J., and Pennebaker, J. Westward. (2011). Linguistic communication style matching predicts relationship initiation and stability. Psychol. Sci. 22, 39–44.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kempen, G., Olsthoorn, N., and Sprenger, S. (2011). Grammatical workspace sharing during language production and language comprehension: evidence from grammatical multitasking. Lang. Cogn. Procedure. 27, 345–380.
Levelt, W. J. K., and Kelter, Southward. (1982). Surface form and memory in question answering. Cogn. Psychol. 14, 78–106.
Markman, A. B., and Makin, V. S. (1998). Referential communication and category acquisition. J. Exp. Psychol. Gen. 127, 331–354.
Pubmed Abstract | Pubmed Full Text
McGettigan, C., Agnew, Z. K., and Scott, S. K. (2010). Are articulatory commands automatically and involuntarily activated during speech perception? Proc. Natl. Acad. Sci. U.s.A. 107, E42–E42.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Newman-Norlund, R. D., Bosga, J., and Meulenbroek, R. G. J. (2008). Anatomical substrates of cooperative joint-action in a continuous motor task: virtual lifting and balancing. Neuroimage 41, 169–177.
Pubmed Abstract | Pubmed Total Text | CrossRef Total Text
Newman-Norlund, R. D., van Schie, H. T., van Zuijlen, A. M. J., and Bekkering, H. (2007). The mirror neuron arrangement is more than agile during complementary compared with imitative action. Nat. Neurosci. x, 817–818.
Pubmed Abstruse | Pubmed Full Text | CrossRef Total Text
Noordzij, M. L., Newman-Norlund, Southward. Eastward., de Ruiter, J. P., Hagoort, P., Levinson, S. C., and Toni, I. (2009). Brain mechanisms underlying man communication. Front. Hum. Neurosci. 3:xiv. doi: 10.3389/neuro.09.014.2009
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Pickering, Chiliad., and Garrod, S. (2006). Alignment as the basis for successful communication. Res. Lang. Comput. 4, 203–228.
Pickering, M. J., and Garrod, S. (in printing). An integrated theory of language production and comprehension. Behav. Brain Sci.
Pulvermüller, F., Huss, Yard., Kherif, F. Moscoso Del Prado Martin, F., Hauk, O., and Shtyrov, Y. (2006). Motor cortex maps articulatory features of speech sounds. Proc. Natl. Acad. Sci. U.s.a.A. 103, 7865–7870.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Redcay, E., Dodell-Feder, D., Pearrow, M. J., Mavros, P. 50., Kleiner, M., Gabrieli, J. D. East., and Saxe, R. (2010). Live face-to-face interaction during fMRI: a new tool for social cognitive neuroscience. Neuroimage 50, 1639–1647.
Pubmed Abstract | Pubmed Total Text | CrossRef Full Text
Richardson, D. C., and Dale, R. (2005). Looking to understand: the coupling between speakers' and listeners' eye movements and its human relationship to discourse comprehension. Cogn. Sci. 29, 1045–1060.
Pubmed Abstruse | Pubmed Total Text | CrossRef Full Text
Rossi, E., Schippers, Yard., and Keysers, C. (2011). "Broca'south area: linking perception and product in linguistic communication and actions," in On Thinking, eds Due south. Han and E. Pöppel (Berlin, Heidelberg: Springer Berlin Heidelberg), 169–184.
Schippers, M. B., Roebroeck, A., Renken, R., Nanetti, Fifty., and Keysers, C. (2010). Mapping the data menses from i brain to another during gestural communication. Proc. Natl. Acad. Sci. UsA. 107, 9388–9393.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Scott, S. K., McGettigan, C., and Eisner, F. (2009). A little more than conversation, a piffling less action: candidate roles for the motor cortex in speech communication perception. Nat. Rev. Neurosci. 10, 295–302.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Segaert, Chiliad., Menenti, L., Weber, Yard., Petersson, K. K., and Hagoort, P. (2011). Shared syntax in language production and language comprehension—an fMRI report. Cereb. Cortex. doi: 10.1093/cercor/bhr249. [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Thompson, C. Grand., Bonakdarpour, B., Set up, Due south. C., Blumenfeld, H. K., Parrish, T. B., Gitelman, D. R., and Mesulam, Chiliad. One thousand. (2007). Neural correlates of verb argument structure processing. J. Cogn. Neurosci. 19, 1753–1767.
Pubmed Abstract | Pubmed Full Text | CrossRef Total Text
Watson, K. Eastward., Pickering, Yard. J., and Branigan, H. P. (2004). "Alignment of reference frames in dialogue," in Proceedings of the 26th Annual Conference of the Cognitive Science Society, (Mahwah, NJ).
Willems, R. M., de Boer, M., de Ruiter, J. P., Noordzij, M. L., Hagoort, P., and Toni, I. (2010). A dissociation betwixt linguistic and chatty abilities in the human brain. Psychol. Sci. 21, 8–14.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Wilson, Yard., and Wilson, T. P. (2005). An oscillator model of the timing of turn-taking. Psychon. Bull. Rev. 12, 957–968.
Pubmed Abstract | Pubmed Full Text
Zwaan, R. A., and Radvansky, K. A. (1998). Situation models in language comprehension and retentiveness. Psychol. Bull. 123, 162–185.
Pubmed Abstract | Pubmed Total Text
Source: https://www.frontiersin.org/articles/10.3389/fnhum.2012.00185/full
0 Response to "Ratusnik Test 1 Focus Review Neuro Basis of Speech and Communication"
Post a Comment